2.3. 线粒体与叶绿体¶
线粒体、叶绿体是真核细胞内两个由双层膜包被的产能细胞器,呈现显著整体性、动态特征;携带遗传物质DNA,以原核编码方式转录合成部分自身所需RNA、蛋白质;经分裂方式增殖;为半自主性细胞器,可能为内共生起源。
2.3.1. 线粒体与氧化磷酸化¶
2.3.1.1. 基本形态与动态特征¶
线粒体(mitochondrion)呈颗粒、短线状,直径0.3~1.0μm、长度1.5~3.0μm;大小、形态可随细胞生命活动变化而变化;胞内能力需求集中区分布密集,时刻处于依赖细胞骨架、马达蛋白的运动中;胞中线粒体数目动态变化、受物种遗传信息调控,与细胞类型相关,随分化而变化。
线粒体处于融合、分裂的动态过程;植物细胞中频繁的融合、分裂可能是线粒体间共享遗传信息的重要途径。融合、分裂过程需大分子GTPase参与。
2.3.1.2. 超微结构¶
线粒体为两层膜包裹的细胞器:外膜(outer membrane)平展,起界膜作用;内膜(inner membrane)内折延伸成嵴(cristae);内外膜间空间为膜间隙(intermembrane space);内膜之内空间为基质(matrix)。
结构 |
特征 |
标志酶 |
功能 |
|---|---|---|---|
外膜 |
平整光滑、通透性高 |
单胺氧化酶 |
膜磷脂合成、底物初步分解 |
膜间隙 |
腺苷酸激酶 |
核苷磷酸化 |
|
内膜 |
无胆固醇、富含心磷脂,通透性低;内折成嵴;含ATP合酶 |
细胞色素氧化酶 |
电子传递、氧化磷酸化、代谢物转运 |
基质 |
具特定pH、渗透压;含TCA、脂肪酸氧化、氨基酸降解相关酶;DNA、RNA、核糖体等 |
苹果酸脱氢酶 |
TCA、脂肪酸氧化、氨基酸降解、蛋白质合成、DNA复制、RNA合成 |
2.3.1.3. 氧化磷酸化¶
线粒体主要功能:高效地将有机物中储存的能量转为细胞生命活动的直接能源ATP;实质是把H+跨膜电位差、H+浓度梯度形成的质子驱动力(proton motive force)转换为ATP分子中的高能磷酸键。
电子传递链(electron transport chain)/呼吸链(respiratory chain):TCA循环的高能电子在线粒体内膜上经各电子载体(electron carrier)有序转移至O2的电子传递序列。
氧化磷酸化(oxidative phosphor-ylation):线粒体中ATP合成与电子传递链的氧化过程偶联。
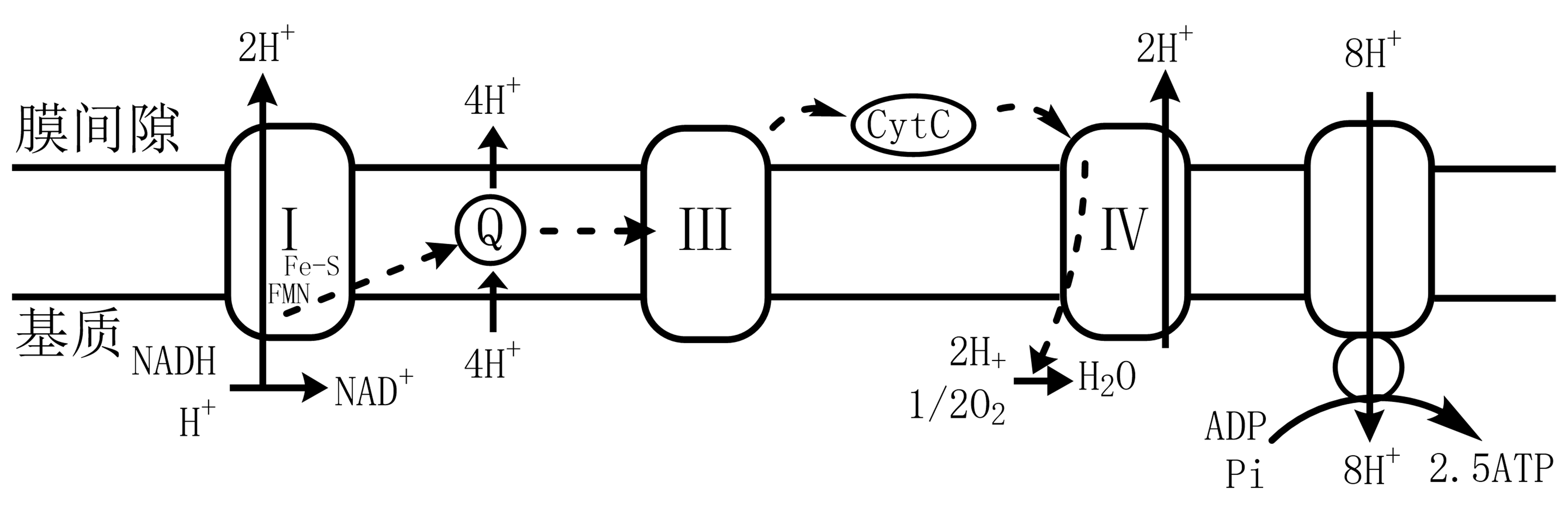
图 2.3.1 电子传递链¶
复合物Ⅰ(NADH-CoQ还原酶)每传递1对电子伴随4个质子从基质转移至膜间隙。
复合体Ⅲ(CoQ-Cyt c还原酶)每传递1对电子时,4个质子转移至膜间隙(其中辅酶Q2个)。
复合体Ⅳ(细胞色素氧化酶)每传递1对电子转移2个质子至膜间隙,2个质子形成水。
2.3.1.4. 疾病¶
线粒体病(mitochondrial disease):由线粒体功能障碍引起的疾病;即可能因线粒体DNA突变,也可能因核DNA突变引发。
2.3.2. 叶绿体与光合作用¶
2.3.2.1. 基本形态与动态特征¶
植物细胞中,叶绿体(chloroplast)为动态细胞器;光调控下分布、位置变化,基质小管介导的相互连接,随分化、去分化的形态变化,分裂导致数目变化等。
叶绿体体积较大、含叶绿素,是最易观察到的细胞器;高等植物叶肉细胞中呈凸透镜状、铁饼状,直径5~10μm、厚2~4μm;叶片平展后叶绿体体积、数目相对保持稳定。
躲避响应(avoidance response):叶绿体经位移避开强光的行为。
积聚响应(accumulation response):较弱光照下叶绿体汇集到细胞受光面。
叶绿体定位(chloroplast positioning):叶绿体于胞内位置、分布受动态调控;移动、锚定借助微丝骨架。
未发育种子(胚)细胞中无叶绿体,萌发时子叶、叶鞘、真叶细胞中原质体(proplastid)分化为叶绿体;此外,储藏组织、其它白化组织中,分化为造粉质体(amyloplast)、白色体(etioplast)。特定条件下叶绿体的分化可逆。
前叶绿体(pre-chloroplast):分化中的叶绿体;幼叶中叶绿体体积较成熟叶绿体小,基质仅形成少数基质类囊体,未形成或正开始形成基粒类囊体。叶绿体数目调控只要发生于细胞分化、生长的早期阶段。
2.3.2.2. 超微结构¶
叶绿体超微结构:叶绿体被膜(chlotoplast envelope)/叶绿体膜(chloroplast membrane)、类囊体(thylakoid)、基质(stroma)。
叶绿体为双层单位膜包被的细胞器,内外膜间腔隙为膜间隙(intermembrane space),外膜通透性大于内膜,内膜上具多种转运蛋白。
类囊体:叶绿体内部由内膜衍生成的封闭扁平膜囊,用以进行光反应。类囊体囊内空间为类囊体腔(thylakoid lumen)。
基粒(grana):叶绿体内圆饼状类囊体有序叠至成垛。
基粒类囊体(granum thylakoid):组成基粒的类囊体。
基质片层(stroma lamella)/基质类囊体(stroma thylakoid):贯穿于基粒间,不形成垛叠的片层结构。
叶绿体基质即内膜与类囊体间的液态胶体物;主要成分为可溶性蛋白、代谢活跃物,丰度最高为核酮糖-1,5-2P羧化酶/加氧酶(Rubiso),还含CO2固定所需酶、叶绿体DNA、脂滴(lipid droplet)、淀粉粒(starch grain)等。
2.3.2.3. 光合作用¶
光合作用是叶绿体主要功能,高等植物中分两步:依赖光的反应(light dependent reaction)/光反应(light reaction)、碳同化反应(carbon assimilation reacton)/固碳反应(carbon fixation reaction)。光反应中含:原初反应、电子传递及光合磷酸化两步。
原初反应(primary reaction):光合色素分子被光能激发引起第一个光化学反应的过程。
光合电子传递链(photosynthetic electron transfer chain)由一系列电子载体构成。
光系统(photosystem,PS):光合作用中光吸收功能单位,由叶绿素、类胡罗卜素、脂质、蛋白质组成;每形成1O2需4e-从2H2O传至2NADP+,共需8光子。
光合磷酸化(photophosphorylation):光照引起电子传递与磷酸化作用偶联生成ATP的过程。
非循环光合磷酸化(noncyclic photophosphorylation):光能驱动电子单向传递。
循环光合磷酸化(cyclic photophosphorylation):光能驱动电子经过PC回至PSⅠ形成闭合回路,仅产生ATP。
二氧化碳同化(CO2 assimilation):光合作用中的固碳反应;卡尔文循环、C4途径、CAM。
2.3.3. 半自主性与起源¶
线粒体、叶绿体的功能主要受核基因组调控,同时受自身基因组调控,故称半自主性细胞器(semiautonomous organelle)。
线粒体DNA(mitochondrial DNA,mtDNA)、叶绿体DNA(cpDNA)均呈环状,对应称线粒体基因组(mitochondrial genome)、叶绿体基因组(chloroplast genome)。
真核细胞中,细胞核与线粒体、叶绿体间遗传信息、基因表达调控层次上建立的分子协调机制称核质互作(nuclear-cytoplasmic interaction),当突变时则称核质冲突(nuclear-cytoplasmic incompatibility,nuclear-cytoplasmic conflict)。
内共生起源(endosymbiotic origin)学说:线粒体、叶绿体起源于原始真核生物内共生的有氧呼吸细菌、光合自养蓝细菌。依据:基因组与细菌基因组具有明显相似性,具完整、独立蛋白质合成系统,分裂方式与细菌类似,膜特性(外膜与真核内膜系统相似性质、内膜与细菌质膜相似);线粒体磷脂、呼吸类型、Cyt c初级结构与某些细菌相近,自然界胞内共生蓝藻具基因片段转移等叶绿体行为特征。