1.7. 氧化、代谢及能学¶
生物氧化:有机分子在胞内氧化分解为CO2、H2O并释放出能量形成ATP的过程;体温进行、由酶催化、逐步氧化放能,产生的能量常贮存于ATP。
1.7.1. 代谢概览¶
新陈代谢:营养物质在生物体内经系列化学变化的总称;为生物体生命活动重要特征之一;由酶催化。代谢中间产物:代谢过程中连续转变的酶促产物。中间代谢:新陈代谢途径中的个别环节、步骤。主要代谢途径:复杂代谢网络中具共同规律的途径。
新陈代谢功能:从环境获得营养物质、将外界营养物质转为自身需要的结构元件、将结构元件装配为自身大分子、形成或分解特殊功能所需的生物分子、提供生命活动所需能量。
分解代谢:有机营养物经系列反应转为较小、较简单物质的过程。分解代谢所经途径即为分解代谢途径,伴随能量释放。合成代谢:利用大小分子的结构元件构建自身大分子的过程;伴随能量供应。通常,同一物质的分解、合成代谢途径不一致,非简单可逆反应,常经不同中间反应或酶实现。两用代谢途径:分解、合成代谢在某些环节上共同利用;TCA。
能量代谢:以物质代谢为基础,与其伴随发生,蕴藏其中的能量转化。太阳能为最根本能量来源。能量传递系统:ATP、ADP及无机磷酸广泛存在,起传递能量的作用。异养生物分解代谢产生ATP的过程:大分子营养物分解为较小分子(无ATP)、各小分子转化为少数共同物质(少量ATP)、共同物质进入TCA代谢(大量ATP)。ATP用途:供给合成所需能量,机体活动、肌肉收缩能量来源,供给营养物逆浓度运输,保证基因信息传递;ATP为递能分子非储能分子。
1.7.2. 辅酶作用¶
NAD、NADP,FMN、FAD为质子、电子传递载体。
CoA分子中含腺嘌呤、D-核糖、磷酸、焦磷酸、泛酸、巯基乙胺。乙酰CoA形成的硫酯键与ATP的高能磷酸键类似;具高乙酰基转移势能,携带为活泼乙酰基团。
1.7.3. 代谢调节及常见机制¶
代谢调节水平:分子水平(反应物、产物调节;酶调节为最基本代谢调节,活性:可逆变构调节、共价修饰,数量:合成速率、降解速率);细胞水平(细胞特殊结构与酶结合,定位、分隔);整体水平(激素、神经调节)。
反应机制:基团转移反应,氧化-还原反应,消除、异构化及重排反应,碳-碳键形成、断裂反应。
1.7.4. 研究方法¶
在体内:in vivo;在体外:in vitro。
酶抑制剂使用;遗传缺陷症研究代谢途径;气体测量;同位素示踪;核磁共振波谱。
1.7.5. 热力学概念与自由能¶
能量关系:\(\Delta G=\Delta H - T\Delta S\);
能量守恒(热力学第一定律):体系及其周围环境的总能量为常数;热力学第二定律:热传导只由高温物体传至低温物体,具方向性。
偶联反应使得一个在热力学上不利的反应可由热力学上有利的反应驱动。
1.7.6. 高能磷酸化合物¶
高能磷酸化合物:磷酰基水解时释放大量自由能;类型:磷氧键型、氮磷键型、硫酯键型、甲硫键型;具高基团转移势能,磷酸肌酸、PEP、乙酰磷酸均高于ATP。
使ATP易于水解的因素:导致反应物不稳定因素、导致产物稳定因素。Glc分解中间物PEP、1,3-BPG,能量经激酶作用转磷酸基至ADP而传递能量。神经、肌肉细胞活动直接功能物为ATP;磷酸精氨酸、磷酸肌酸以高能磷酸基团为贮能物(磷酸原)。
ATP动态平衡:胞内调节机制,即调节ATP生成以供给能量,也调节ATP利用维持相对稳定。ATP分子中与核糖相连的α-磷酸基团周转率很低。
\(能荷=\frac{[ATP] + \frac{1}{2}[ADP]}{[ATP]+[ADP]+[AMP]}\);
\(磷酸化势能=\frac{[ATP]}{[ADP][Pi]}\);
1.7.7. 电子传递链与氧化磷酸化¶
生物氧化实质为氧化磷酸化,即NADH、FADH2上的电子经系列电子传递体至O2,伴随NADH、FADH2在氧化,释放能量合成ATP。
1.7.7.1. 电子传递链¶
呼吸链(电子传递链):电子从NADH至O2的过程。
抑制剂:
复合体Ⅰ:鱼藤酮、安密妥;
复合体Ⅲ:抗霉素A;
复合体Ⅳ:CN-、N2-、CO。
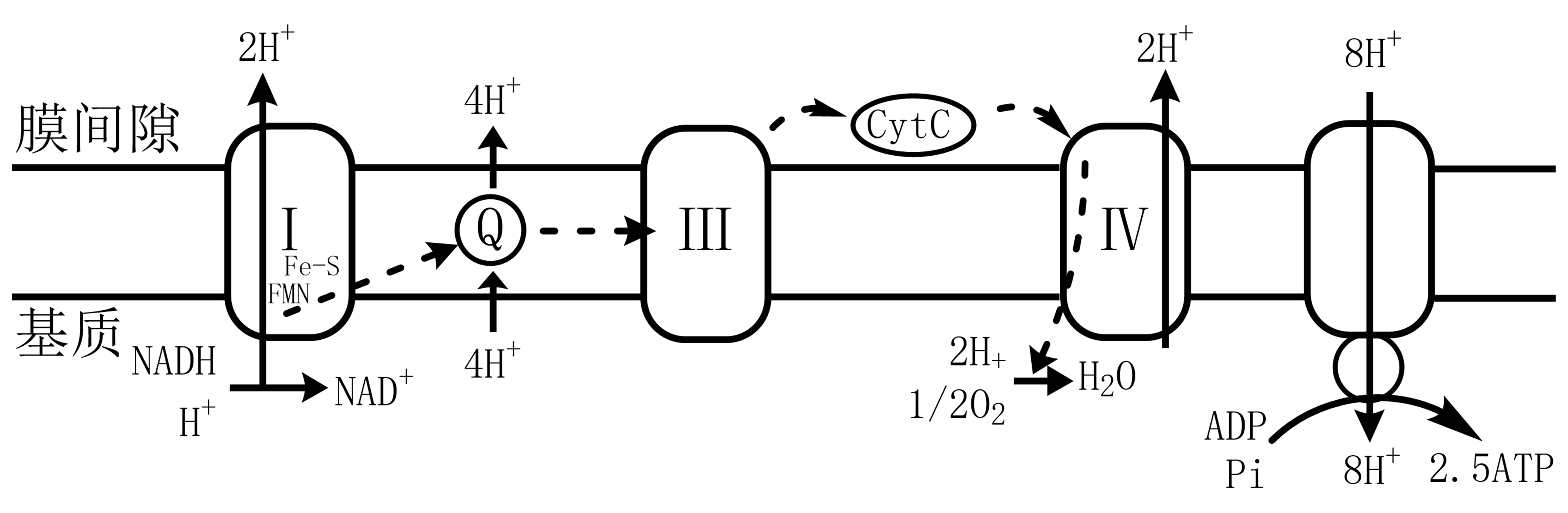
图 1.7.1 线粒体电子传递链及氧化磷酸化¶
1.7.7.2. 氧化磷酸化¶
氧化磷酸化:需氧细胞生命活动的主要能量来源、生物合成ATP主要途径;直接与电子传递链偶联的由ADP形成ATP的磷酸化作用。
底物水平磷酸化:ATP的形成直接由代谢中间物磷酸基团转移至ADP,如PEP。
ATP合成由ATP合酶(复合体Ⅴ)合成,能量偶联(化学偶联假说、构象偶联假说、化学渗透假说)。
调节:ADP为关键物质,调节氧化磷酸化作用(呼吸控制)。
化学渗透假说:电子传递链释放的自由能及ATP合成与跨线粒体内膜的质子梯度偶联;即电子传递的自由能驱动H+从线粒体基质跨内膜至膜间隙,形成H+电化学梯度;要点:
氧化磷酸化需封闭的线粒体内膜,
线粒体内膜对H+、OH-、K+、Cl-等不通透,
破坏H+浓度梯度形成必然破坏氧化磷酸化进行,
线粒体电子传递形成的电子流可将H+从膜内驱动至膜间隙,
大量直接、间接实验证据表明膜表面可滞留大量质子,一定条件下质子沿膜表面迅速移动、远超水相中移动速度。
氧化磷酸化解偶联剂:使电子传递与ATP形成过程分离;仅抑制ATP形成、不抑制电子传递过程,电子传递产生的自由能转为热能;DNP增加内膜H+通透性。
抑制剂:
抑制ATP形成、氧利用,不直接抑制电子传递链载体;
寡霉素,可被DNP解除抑制。
离子载体抑制剂:增加内膜对阳离子通透性破坏氧化磷酸化。
1.7.7.3. 穿梭系统¶
胞质中的NADH不能透过线粒体内膜进入线粒体氧化,需要穿梭系统将其运输至线粒体内。甘油-3-磷酸穿梭途径、苹果酸-天冬氨酸穿梭途径。
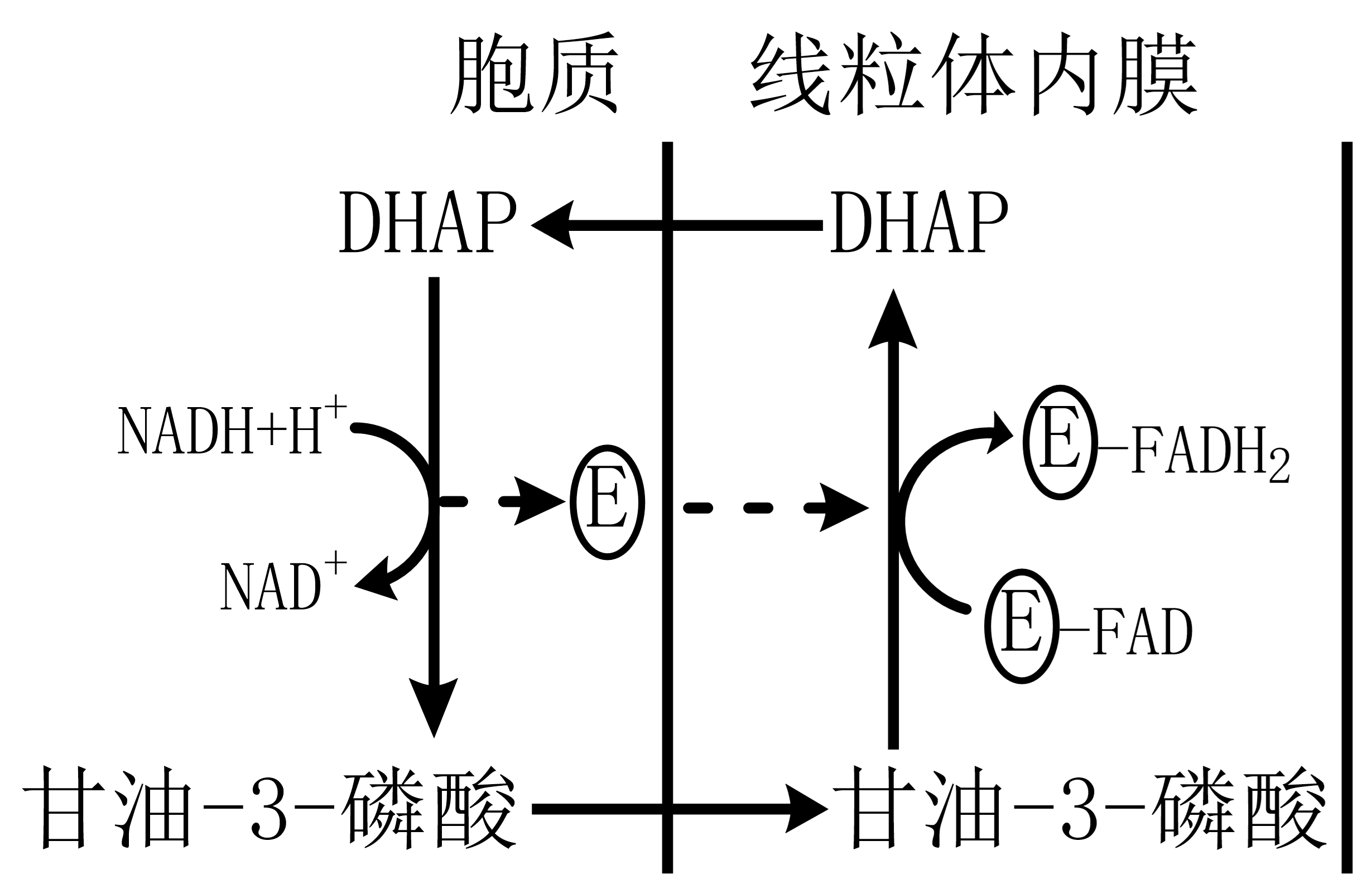
图 1.7.2 甘油-3-磷酸穿梭系统¶
甘油-3-磷酸穿梭系统:胞质中NADH上的电子经穿梭运输后转至FADH2,使得最终ATP产量为1.5非2.5;意义在于:使胞质中NADH逆浓度运输至线粒体内膜,进入电子传递链氧化。昆虫飞行肌明显。
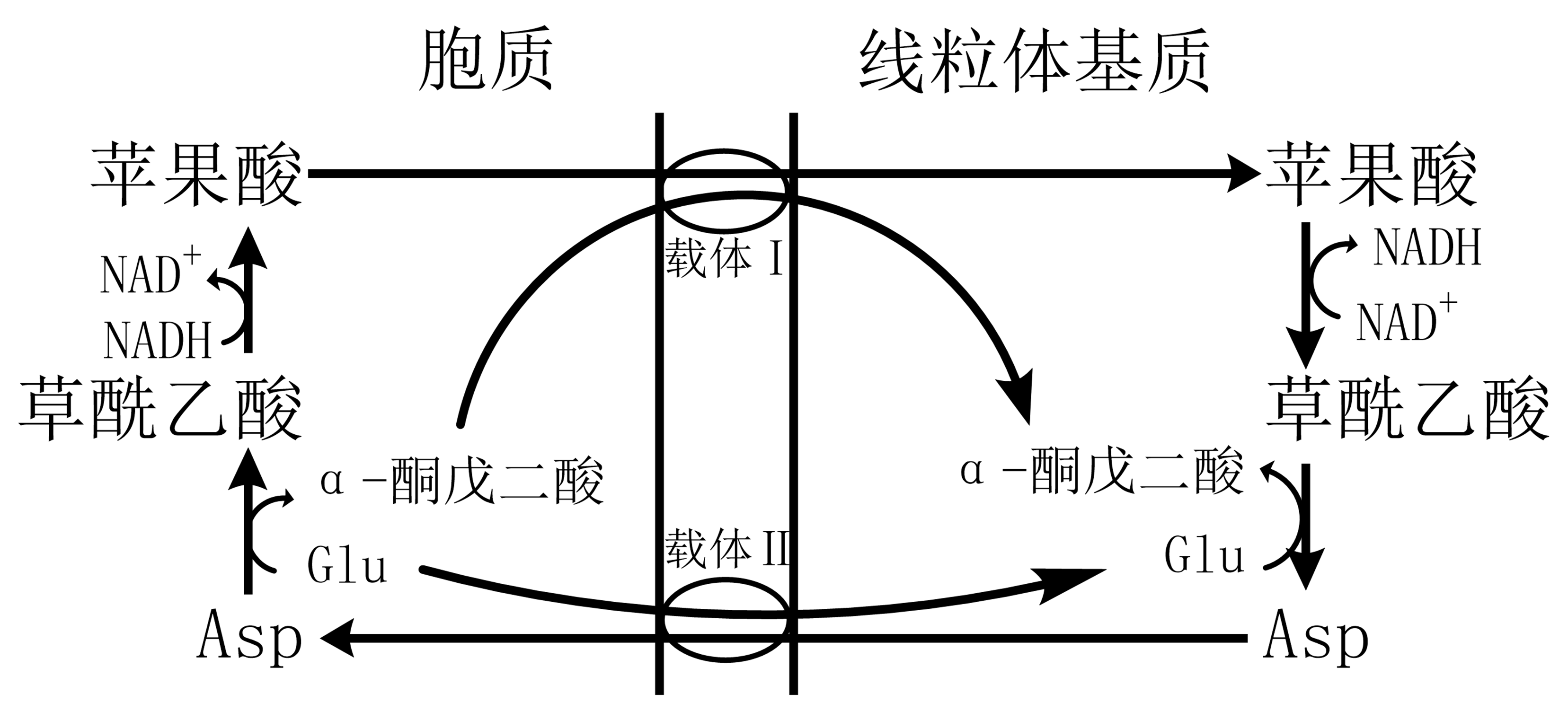
图 1.7.3 苹果酸-天冬氨酸穿梭系统¶
苹果酸-天冬氨酸穿梭:具可逆性,仅当胞质中NADH/NAD+高于线粒体基质时,NADH才经此进入线粒体;( 图 1.7.3 中载体Ⅰ:苹果酸-α-酮戊二酸载体;载体Ⅱ:Glu-Asp载体。)。
1.7.8. Glc彻底氧化生成ATP量¶
反应 |
ATP量 |
|---|---|
糖酵解 |
|
Glc磷酸化 |
-1 |
F6P磷酸化 |
-1 |
2 GAP氧化产2 NADH(①) |
|
2 1,3-BPG去磷酸化 |
+2 |
2 PEP去磷酸化 |
+2 |
丙酮酸转乙酰CoA产2 NADH(②) |
|
TCA |
|
2 异柠檬酸氧化产2 NADH(③) |
|
2 α-酮戊二酸氧化产2 NADH(④) |
|
2 琥珀酰CoA产2 GTP |
+2 |
2 琥珀酸氧化产2 FADH2(⑤) |
|
2 苹果酸氧化产2 NADH(⑥) |
|
氧化磷酸化 |
|
①中2 NADH(设经甘油磷酸穿梭) |
+3 |
②中2 NADH(发生于线粒体内) |
+5 |
③、④、⑥共6 NADH |
+15 |
⑤中2 FADH2 |
+3 |
总计 |
+30 |