1.1. 糖类及其代谢¶
1.1.1. 概述¶
糖类:多羟醛、多羟酮或其衍生物,或水解时能产生这类化合物的物质。糖类物质干重占植物85%-90%,细菌10%-30%,动物小于2%。根本来源是绿色细胞进行的光合作用。
1.1.1.1. 生物学作用¶
作为生物体的**结构成分**;
作为生物体内的**主要能源物质**;
在生物体内转变为其它物质;
作为细胞识别的**信息分子**。
1.1.1.2. 命名分类¶
单糖:不能被水解成更小分子的糖类(葡、果、核)。
寡糖:双糖(麦、蔗)、三糖(棉子糖)、四糖等,20残基以下。
多糖:水解产生20个单糖以上。
同多糖:水解产生同种单糖或其衍生物(糖原、淀粉、壳多糖)。
杂多糖:水解产生不止一种单糖(透明质酸、半纤维素)。
糖复合物:糖类与蛋白质、脂质等生物分子形成的共价结合物(糖蛋白、蛋白聚糖、糖脂)。
1.1.1.3. 旋光异构¶
同分异构:指存在两个或多个具有相同数目和种类的原子而具有相同分子质量的化合物(组成):
结构异构(分子中原子次序不同,构造)
立体异构(原子在空间的相对分布或排列,构型)
几何异构(顺反异构,双键或环等限制原子间自旋)
旋光异构(分子内存在不对称碳原子,手性分子)。
构象:分子所采取的特定形态;围绕单键旋转引起组成原子的不同排列。
平面偏振光(光经Nicol棱镜后,因仅允许某一偏振面光波通过而成)通过旋光物质溶液时使光偏振面向右(+)或左(-)旋转。一定条件下,旋光度(α)与待测液浓度(c)和通过路径长度(l)的乘积成正比。
任一旋光化合物都只有一个对映体,其余为非对映(异构)体(化学和物理性质等不同)。仅一个手性碳原子的构型不同的非对映异构体称为差向异构体。
不对称碳原子(手性碳原子、手性中心、不对称中心,C*)指与四个不同原子或基团共价连接并失去对称性的四面体碳。N个手性碳原子具有2n个旋光异构数及2n-1对对应体。
Fischer投影式,立体模型或透视结构在纸面上的投影。直线交叉点为碳原子,水平方向键向纸面前方,垂直方向键向纸面后方。
RS表示法,先确定取代基优先顺序(顺序规则:原子序数高为优先),后使序数最低的远离余下三个取代基的顺序为顺时针为R构型(+),逆时针为S构型(-)。
1.1.2. 单糖¶
Glc与乙酸酐加热形成无乙酸酯结晶(Glc含5个羟基),与无水氰化氢加成的氰醇衍生物水解碘酸还原得正庚酸(Glc为直链己醛)。
Fru经上述步骤得无乙酸酯和2-甲基己酸(己酮糖)。
D系醛糖由D-甘油醛衍生,L系醛糖由L-甘油醛衍生。糖的构型(D、L)与旋光方向(+、-)无直接关系。
变旋现象:许多单糖新配置溶液会发生旋光度改变。
Glc(多羟基醛)不能与Schiff试剂(品红-亚硫酸)发生紫色反应、难与亚硫酸氢钠发生加成反应、在无水甲醇中经氯化氢催化得含一个甲基化合物(表现缩醛特征:碱稳定、无还原性、无羰基反应及易被酸水解)。
单糖由直链变为环状结构后,羰基碳成为新手性中心,导致C1差向异构化形成两个非对映异构体,羰基碳上形成的差向异构体称异头物。异头碳(半缩醛碳)羟基与最末手性碳的羟基相同取向为α,相反为β。
开链单糖成环状半缩醛时易形成五元环(呋喃:椅式、船式构象)或六元环(吡喃)。异头碳羟基与末端羟甲基反式为α、顺式为β。
1.1.2.1. 单糖性质¶
物理性质:旋光性、甜度、溶解度(微溶于乙醇、不溶于乙醚、丙酮等非极性有机溶剂)。
化学性质:异构化(弱碱作用):室温碱催化的烯醇化作用结果。
氧化:
成醛糖酸:游离醛基具还原性,碱性溶液中重金属离子使醛糖醛基氧化成羧基。如:Fehling试剂和Benedict试剂(较稳定不易受肌酸尿酸等干扰常用于尿糖定性半定量测试)。缓冲的溴水溶液专一氧化醛糖为一羧酸。(还原糖:所有醛糖、许多酮糖。)
成醛糖二酸:醛糖醛基和伯醇基在较强氧化剂(稀硝酸)下均氧化成羧基成二羧酸。单糖氧化成糖二酸测定旋光性对推断单糖手性碳构型具重要意义(半乳糖氧化产物乳糖二酸内消旋)。
成糖醛酸:某些醛糖在特定脱氢酶作用下只氧化伯醇基保留醛基成糖醛酸。
还原:单糖羰基在还原条件下(如硼氢化钠)可还原成多元醇(糖醇)。
成糖脎:苯肼与还原糖反应所成含两个苯腙基衍生物(只与醛基侧两个碳相关,其余碳无关)。
成脂:碱催化下酰氯或酸酐反应,所有羟基含异头碳羟基都酯化。
成醚:在甲基亚磺酰甲基钠下用碘甲烷或碱性条件下用硫酸二甲酯可得糖或糖苷的甲醚衍生物(糖的甲基化)。五甲基Glc上,缩醛甲氧基可通过酸水解去除。
成糖苷:环状单糖的半缩醛或半缩酮羟基与另一化合物缩合成的缩醛或缩酮称糖苷或苷。
单糖脱水:戊糖与12%HCl共热脱水生成糠醛;己糖与酸共热成5-羟甲糠醛(比糠醛易溶于酸),可分解为乙酰丙酸、甲酸及暗色不溶物(腐黑物)。糠醛与多元酚产生特有颜色反应。 表 1.1.1
糖的高碘酸化:高碘酸及其盐(IO4-)可定量氧化断裂邻二羟基、α-羟基醛等碳碳键。可根据消耗的IO4-量或生成甲酸量确定为呋喃糖(1IO4-、0甲酸)或吡喃糖(2IO4-、1甲酸)。
单糖链延长及缩短:Kiliani-Fischer合成,醛糖醛基转为氰醇腈基再转为延长后的醛糖。Wohl降解,醛糖醛式羰基转为腈基后形成氰醇,在碱性条件下脱去HCN转为链缩短1个碳的醛糖。
糠醛、多元酚 |
颜色反应 |
备注 |
|---|---|---|
羟甲糠醛、间苯二酚 |
红色 |
|
糠醛(戊糖)、间苯三酚 |
朱红色 |
间苯三酚实验 |
糠醛(戊糖)、甲基间苯二酚 |
蓝绿色或橄榄绿色 |
|
糠醛及其衍生物(糖)、α-萘酚 |
红紫色 |
|
糖类脱水物、蒽酮 |
蓝绿色 |
|
1.1.3. 重要单糖及衍生物¶
1.1.3.1. 单糖¶
丙糖:D-甘油醛(具光学活性的最简单单糖)、二羟丙酮(无光学活性),其磷酸酯为糖酵解中间物。
丁糖:D-赤藓糖(其磷酸酯为戊糖磷酸途径及Calvin循环中间物)、D-赤藓酮糖,常见于藻类、地衣等低等植物中。
戊糖:D-核糖(RNA、其磷酸酯为戊糖磷酸途径及Calvin循环中间物)、2-脱氧-D-核糖(DNA)、D-木糖(以多聚戊糖形式存在于植物、细菌细胞壁,树胶、半纤维素组分)、L-阿拉伯糖(果胶糖,为果胶物质、半纤维素、树胶、植物糖蛋白的重要组分)、D-阿拉伯糖(植物糖苷、细胞壁组成)、芹菜唐(支链戊糖,D-赤藓呋喃糖形式存在)、D-核酮糖及D-木酮糖(其磷酸酯为戊糖磷酸途径及Calvin循环中间物)。
己糖:D-葡萄糖、D-半乳糖(乳糖、蜜二糖、棉子糖、某些糖苷及脑苷脂、神经节苷脂组分)、L-半乳糖(构件分子,琼脂)、D-甘露糖(甘露聚糖形式于植物细胞壁)、D-果糖、L-山梨糖(工业合成维生素C重要中间物)。
庚糖、辛糖:D-景天庚酮糖(其磷酸酯为戊糖磷酸途径及Calvin循环中间物)、D-甘露庚酮糖、L-甘油-D-甘露庚糖(沙门氏菌细胞壁外层脂多糖结构组分)、D-甘油-D-甘露辛酮糖(和D-甘露庚酮糖不能被人体代谢,尿糖反应阳性)。
1.1.3.2. 单糖磷酸酯¶
单糖磷酸酯(磷酸化单糖)广泛存在于各种细胞中,为代谢途径中主要参与者。
糖的磷酸酯酸性比H2PO4强,再细胞内以1价、2价阴离子混合物形式存在。磷酸酯的水解是P-O键断裂引起。
成线机制:磷酸酯水解及磷酸基团的转移时,进入基团(RO-\HO-)在离去基团的对面向磷原子亲核攻击,形成五价过渡态,脱去离去基团;在三角双椎体中进入基团与离去基团共线。
1.1.3.3. 糖醇¶
单糖羰基被还原生成糖醇。
山梨醇(D-葡萄醇,糖尿病患者眼球晶状体山梨醇累积导致白内障)
D-甘露醇(降低颅内压、治疗急性肾功能衰竭)
半乳糖醇(半乳糖还原产物)
核糖醇(参与形成核黄素VB2)
木糖醇(G6P经糖醛酸途径代谢中间物)
肌醇:环己六醇,9个立体异构体,7个为内消旋体:顺、表、别、新、肌、粘、鲨,一对对映体:1L-手性、1D-手性
鲨肌醇最稳定
肌肌醇:唯一活性异构体;酵母、白鼠等重要生长因子VB,对人等某些动物为非必需维生素;参与部分磷脂组成、第二信使。
1.1.3.4. 糖酸¶
醛糖氧化条件不同可被氧化成3类糖酸(糖羧酸):醛糖酸(内酯)、糖二酸(少见、广泛存于植物界)、糖醛酸(内酯)。
生物体内无游离醛糖酸,有其衍生物参与代谢等。葡糖酸可与钙、铁等离子形成可溶性盐易被吸收;葡糖酸钙常用于治疗钙缺乏及过敏性疾病。抗坏血酸(VC),醛糖酸衍生物,酸性来自C3上烯醇式羟基氢解离,
1.1.3.5. 脱氧糖¶
脱氧糖指分子中一个或多个羟基被氢原子取代的糖。广泛分布于植物、细菌及动物中。L-鼠李糖、L-岩藻糖、D(+)-毛地黄毒素糖、泊雷糖、阿比可糖、泰威糖。
1.1.3.6. 氨基糖¶
氨基糖是分子中一个羟基被氨基取代的单糖。氨基多以乙酰氨基形式存在,少数游离。如:葡糖胺、N-乙酰葡糖胺、半乳糖胺、N-乙酰半乳糖胺。胞壁酸和神经氨酸称酸性氨基糖(酸性糖)。N-乙酰胞壁酸是细菌细胞壁结构多糖构件分子之一,由N-乙酰-D-葡糖胺C3上羟基与D-乳酸羟基成醚键连接。神经氨酸,含一个氨基的9C糖酸,骨架由丙酮酸和D甘露糖氨构成。 表 1.1.2
单糖 |
单糖 |
||
|---|---|---|---|
阿拉伯糖 |
Ara |
葡糖酸 |
GlcA |
果糖 |
Fru |
葡糖醛酸 |
GlcUA |
岩藻糖 |
Fuc |
半乳糖胺 |
GalN |
半乳糖 |
Gal |
葡糖胺 |
GlcN |
葡萄糖 |
Glc |
N-乙酰半乳糖胺 |
GalNAc |
来苏糖 |
Lyx |
N-乙酰葡糖胺 |
GlcNAc |
甘露糖 |
Man |
胞壁酸 |
Mur |
鼠李糖 |
Rha |
N-乙酰胞壁酸 |
MurNAc |
核糖 |
Rib |
N-乙酰神经氨酸 |
NeuNAc |
木糖 |
Xyl |
唾液酸 |
Sia |
1.1.3.7. 糖苷¶
苦杏仁苷:龙胆二糖基、氢氰酸苯甲酸残基,β糖苷键;β-葡糖苷酶水解为2Glc、1苯甲酸、1HCN;人体大量摄入引起氢氰酸中毒
毛地黄毒苷:3个D-毛地黄毒素糖经β-1,4糖苷键相连成糖基、毛地黄毒苷配基;强心苷之一,强心苷可治疗充血性心衰及节律障碍,生理活性主要由配基决定
乌本苷:L-Rha糖基、乌本苷配基;Na+、K+-ATP酶强抑制剂
黑芥子硫苷酸钾(S-苷类)
橘皮苷及芸香苷:VP,维持血管正常功能,改善微血管脆性和通透性
根皮苷:可阻断肾小管对Glc重吸收
皂苷:寡糖基、类固醇及多环三萜配基;去污剂
花色素苷
1.1.4. 寡糖¶
寡糖由2~20个单糖经糖苷键连接而成。与多糖无绝对界限,常与聚糖同义。
初生寡糖,在生物体内有相当量,游离存在。
次生寡糖,结构复杂的高级糖,主要作为结构成分。
1.1.4.1. 结构与性质¶
不同的寡糖间主要有4项不同:
参与组成单糖单位
形成糖苷键类型
成键的异头碳羟基构型(异头碳构型对寡糖分子现状影响很大,异头定向)
单糖单位次序(还原端RE、非还原端NRE)
命名:寡糖非还原端于左侧;在首个单糖单位名前加O,示糖单位经氧原子相连;给出异头碳构型;在糖单位名称中表示吡喃(p)或呋喃(f);标示糖苷键类型。
麦芽糖[O-α-D-吡喃葡糖基-(1→4)-β-D-吡喃葡糖]或[O-α-D-Glcp-(1→4)-β-D-Glcp]或[Glcα(1→4)Glc]
1.1.4.2. 常见二塘¶
蔗糖,Glc(α1↔β2)Fru;无Fehling反应、不成脎、无变旋现象,可被α-葡糖苷酶、蔗糖酶(转化酶、β-呋喃果糖苷酶);蔗糖水解时比旋由正转负(转化);溶解度大,大多数生物活性不受高浓度蔗糖影响,适于植物组织间糖运输;水解自由能比淀粉α(1→4)糖苷键自由能高;酸性溶液中极易水解。
乳糖,Galβ(1→4)Glc;具还原性、能成脎、具变旋现象。
麦芽糖,Glcα(1→4)Glc;还原糖。
α,α-海藻糖,Glc(α1↔α1)Glc;非还原糖,初生寡糖;在蕨类中替代蔗糖为主要可溶性储存糖类,昆虫中为主要血循环糖。
纤维二糖,Glcβ(1→4)Glc;次生寡糖,糖环中所有羟基及羟甲基均处于平伏位。
龙胆二糖,Glcβ(1→6)Glc;具还原性、具变旋现象。
1.1.4.3. 其它简单寡糖¶
三糖:
棉子糖:Galα(1→6)Glcα(1→2)Fru;非还原糖;α-半乳糖苷酶水解产生半乳糖和蔗糖,蔗糖酶水解产生果糖和蜜二糖
龙胆糖:Glcβ(1→6)Glc(α1↔β2)Fru;非还原糖
龙胆三糖:还原糖,Glcβ(1→6)Glcβ(1→6)Glc
松三糖:Galα(1→3)Fru(β2 ↔α1)Glc;非还原糖
水苏糖:四糖;棉子糖系列,在棉子糖基础上Gal通过α(1→6)连接到棉子糖Gal上
环糊精:某些芽孢杆菌中的环糊精葡糖基转移酶作用于淀粉(直链为佳)后形成
对酸水解较慢,对α-、β-淀粉酶有较大抗性
分子以单体堆叠成圆筒状多聚体,内部疏水、外部亲水,可形成水溶性包含络合物;可作为稳定剂、抗氧化剂、抗光解剂、乳化剂及增溶剂等
α-环糊精用于层析分离和光谱学测定,β-环糊精于丹磺酰氯形成水溶性笼形物用于蛋白质荧光标记
α-环糊精能使苯酯水解速度增加300倍,β-环糊精能使磷酸酯水解速度增加200+倍,是研究模拟酶材料
1.1.5. 多糖¶
多糖(聚糖),由很多单糖单位构成。相对分子质量极大,大多不溶于水,降解不均一。属非还原糖,无变旋现象,无甜味,常不结晶。
单糖单位来源分:植物多糖、动物多糖、微生物多糖;
单糖单位组成种类分:同多糖、杂多糖;
生物学功能分:贮存多糖(淀粉、糖原、右旋糖苷、菊粉)、结构多糖(纤维素、壳多糖、许多植物多糖、细菌杂多糖、动物杂多糖)。
1.1.5.1. 同多糖¶
淀粉,植物生长时以淀粉粒贮存于细胞种的贮存多糖,富含于种子、块茎、块根。
天然淀粉:
直链淀粉:淀粉胶悬液用微溶于水的醇饱和时的微晶沉淀;微溶于水;由Glc以α-1,4连接;呈左手螺旋,6残基每圈、螺距0.8nm、直径1.4nm;遇碘呈深蓝色,约需36个/6圈Glc
支链淀粉:用于水混溶的醇得的无定形物质;易溶于水;约25至30单位1分支点,该处由Glc以α-1,6连接;其余直链处以α-1,4连接;遇碘呈紫色到紫红色
糊精,淀粉在酸或酶作用下逐步降解形成的分子大小不一的中间物;依分子量递减与碘反应由蓝紫色、紫色、红色至无色。
α-淀粉酶,广泛存于动植物及微生物中,内切葡糖苷酶,随机作用于α-1,4糖苷键(断开C1-O),终产物:葡萄糖、麦芽糖、麦芽三糖及α-(极限)糊精。α-糊精含α-1,6键,继续降解需寡-1,6-葡糖苷酶(异麦芽糖酶/α-糊精酶)。
β-淀粉酶,存于高等植物特别是发芽种子中,外切葡糖苷酶,从非还原端始断裂α-1,4糖苷键,切除二塘单位(β-麦芽糖),不能断裂α-1,6糖苷键,余下大分子核心(β-极限糊精,能被异淀粉酶除去α-1,6糖苷键后继续水解)。
糖原:动物淀粉,颗粒形式存于动物细胞液内,主要存于肝脏及骨骼肌。
备注
支链淀粉和糖原的高度分支可以增加分子溶解度,具更多非还原端利于降解作用,加速聚合物转化为单体,利于葡萄糖的即时动员。
直链淀粉主要用作葡萄糖的长期贮存。
动员主要由(淀粉/糖原)磷酸化酶作用。
改型淀粉:为特定需要将天然淀粉经适当处理改变某些物化性质。
凝胶化(糊化):干淀粉悬于水中加热时,淀粉粒吸水膨胀破裂,淀粉分子进入水中成半透明胶悬液,失去晶态及双折射性质。
退行(老化),凝胶化的淀粉液冷却久置时,淀粉分子聚集并由分子间氢键键合成不溶性微晶束沉淀。
右旋糖酐,细菌及酵母的贮存多糖,主链α-1,6Glc,支链α-1,3Glc或异麦芽糖基。可用作血浆代用品、牙斑主要成分。
菊粉,一种果聚糖,在很多植物种替代淀粉为贮存多糖,约31个β-Fruf及1-2个Glcp聚合而成。Fru间以β(2→1)连接,一个Glc以蔗糖型(α1↔β2)连接,余下Glc若有则位于链内。人及动物缺乏相应酶,可用于测定肾功能。
纤维素,β(1→4)糖苷键。胶束(微晶),若干条链聚集成有周期性晶格的分子束。微纤维,电镜下多个胶束平行呈线状,胶束间具疏松无序多糖链;由纤维素合成酶催化
壳多糖(几丁质):N-乙酰-β-D-葡糖胺的同聚物。
1.1.5.2. 杂多糖¶
果胶物质:主要存于植物初生细胞壁和细胞间的中层内;可从植物材料经乙醇:苯(9:1)脱脂及亚氯酸去木质素后用螯合剂水溶液提取获得;含酸性多糖(聚半乳糖醛酸、聚鼠李乳糖醛酸)、中性多糖(阿拉伯聚糖、半乳聚糖、阿拉伯半乳聚糖)。
果胶:羧基不同程度被甲酯化的线性聚半乳糖醛酸或聚鼠李乳糖醛酸。
果胶酸:完全去甲酯化的果胶。
果胶酸酯:羧基大部分被甲酯化的果胶。
原果胶:存于植物中与纤维素及半纤维素等结合的水不溶性果胶物质。
果胶经果胶酯酶去甲酯化成无黏性果胶酸。
半纤维素:碱溶性植物细胞壁多糖,去除果胶物质后能被15%NaOH提取的多糖。含木聚糖、葡甘露聚糖和半乳葡甘露聚糖、木葡聚糖。
琼脂(洋菜):从红藻类石花菜属及其它属的某些海藻中提取的多糖混合物。含琼脂糖(平行链左手螺旋,三重螺轴,螺距1.9nm)、琼脂胶。多种具相同主链但不同程度被负电基团取代的多糖混合物。
角叉聚糖(卡拉胶):右手双螺旋,3个二糖单元每圈,螺距2.6nm。
藻酸或褐藻酸:褐藻类中部分海藻含有。可用于果胶及琼脂代用品。
树胶或胶质:化学上类似于半纤维素,但更复杂;常不与纤维素集合,能溶于水成极粘稠溶液。
阿拉伯胶、西黄芪胶(黄芪胶)、瓜尔豆胶、黄杆胶(黄原胶)。
1.1.5.3. 细菌多糖¶
细菌细胞壁:主要由多糖组成含脂质及蛋白质;依结构不同,可分为革兰氏阳性菌细胞壁(多层网状结构肽聚糖组成,有磷壁酸相连)和革兰氏阴性菌细胞壁(单层肽聚糖无磷壁酸,外覆脂双层外膜)。
肽聚糖(黏肽、胞壁质):壳多糖链单糖残基交替被乳酸取代,并连接四肽侧链。G-中四肽侧链直接相连;G+中四肽侧链经肽桥连接。
磷壁酸:主链由醇(核糖醇/甘油)及磷酸分子交替连接,侧链为单个Ala或Glc;可调节正离子进出细胞、影响细胞生长(调节自溶素活性,维持细胞壁稳定)、参与磷存储、构成细胞壁抗原特异性。
脂多糖(内毒素):G-特有结构成分,外膜外表面主要物质,亲水,对许多疏水物质起通透性屏障;能引起多种生物效应,内毒活性。由脂质A和杂多糖(核心寡糖、O-特异链)共价连接构成。
荚膜多糖:病原菌具毒性原因。
1.1.6. 糖蛋白及其糖链¶
糖蛋白是一类复合糖或一类缀合蛋白质,糖链为缀合蛋白质的辅基。
1.1.6.1. 糖蛋白及糖链结构¶
许多膜蛋白(血型抗原、组织相容性抗原、移植抗原、膜受体等)及分泌蛋白都是糖蛋白。
不同糖蛋白含糖量变化大,糖可沿肽链均匀分布或集中特定区域。
寡糖链中单糖种类、连接位置、糖苷键构型、糖环类型及支链等构成糖链结构的多样性。
1.1.6.2. 糖肽键类型与糖链分类¶
糖蛋白中寡糖链还原端残基与肽链氨基酸残基共价连接(糖肽键),主要类型:N-糖肽键、O-糖肽键。
N-糖肽键:β构型的N-乙酰葡糖胺异头碳与Asn的γ酰胺N原子共价连接形成。分布广,特别是血浆蛋白及膜蛋白。Asn于肽段Asn-X(非Pro)-Thr/Ser中,GlcNAc-Asn对弱碱稳定,在1MNaOH下易水解。
O-糖肽键:单糖异头碳与羟基氨基酸羟基O共价结合。
GalNAc与Ser/Thr型:单糖GlcNAc、Gal、Man、Xyl、Ara也可参与;粘蛋白型糖肽键;对碱不稳定,易β消去
Gal与Hyl型:存在于胶原蛋白,对碱稳定
Araf与Hyp型:对碱稳定
Cys型:Gal/Glc-Cys
Asp:侧链羧基参与型
糖链的分类依照糖肽键类型可分为 N-连接糖链 及 O-连接糖链。可单独或同时出现于同一糖蛋白中。
N-糖链:含共同结构花式(核心五糖,三甘露糖基核心;进入糖蛋白前以前体G寡糖形式合成)。可分三类:
复杂型:除三甘露糖基核心外无Man;结构变化大:外链数目可多至5、结构多种多样;常含Fuc及GlcNAc
高甘露糖型:除核心五糖外只含Man,具两分支七糖结构共同核心
杂合型:具复杂型及高甘露糖型结构
O-糖链:结构较N-糖链简单,连接形式更多。无共同核心结构。
1.1.6.3. 糖链生物学功能¶
糖蛋白的糖链参与肽链折叠缔合,参与糖蛋白转运分泌,参与分子识别及细胞识别。
N-糖链参与新生肽链折叠,维持蛋白质正确构象。G寡糖中Glc与肽链折叠关系密切。与亚基缔合的作用可能为维持亚基正确构象及亚基间通过糖链相互识别而发生缔合。
分子识别指生物分子的选择性相互作用,是通过两分子的结合部位实现,需结合部位是结构互补、有相应基团相互产生足够作用力。细胞识别实际上是细胞表面分子的相互识别。
受体是位于细胞膜上、细胞质或细胞核中能与胞外生物活性分子专一结合并将信息传递至效应器引起相应生物学效应的生物大分子。配体是被受体识别并结合的生物活性分子。
血清中较多蛋白含以Sia为末端的N-糖链糖蛋白(唾液酸糖蛋白),在血液循环时Sia被切除暴露出Gal,最终被干细胞受体集合后清除。
细胞黏着,多细胞生物中细胞相互识别而聚集成细胞群。胞间充满胞外基质,黏着通过相关膜内在蛋白(细胞黏着分子)介导。细胞黏着分子多数含N-糖链。
淋巴归巢,淋巴细胞迁移至派生部位的现象。依赖归巢受体及血管地址素间的相互识别。
1.1.6.4. 糖链与糖蛋白的生物活性¶
酶活性相关:糖链在酶新生肽链折叠、转运、保护等起作用,但与成熟酶活性关系因酶而异。
激素活性:
IgG活性:糖链参与IgG构成,N-糖链缺失外链Gal后形成自身抗原,引起自身免疫病。
1.1.6.5. 血型及凝集素¶
凝集原:红细胞膜中提取的血型抗原。凝集原的血型决定簇是寡糖。
ABO血型系统:
血型A含凝集原A
血型B含凝集原B
血型AB含凝集原A及B
血型O不含凝集原A及B。
红细胞凝集素,凝集原抗体。抗原系统为ABH。A抗原:GalNAc,B抗原:Gal。
凝集素:一类非抗体蛋白质或糖蛋白能与糖类专一的非共价结合并具有凝集细胞及沉淀聚糖和复合糖作用。
1.1.7. 糖胺聚糖和蛋白聚糖¶
糖胺聚糖和蛋白聚糖是细胞外基质重要成分。
1.1.7.1. 糖胺聚糖¶
糖胺聚糖(粘多糖、氨基多糖、酸性多糖):是动植物特别是高等动物结缔组织中的一类结构多糖。属杂多糖,不分支长链聚合物,由重复二糖单位构成。
生物学作用:保持疏松结缔组织水分,调节阳离子在组织中的分布,对关节的润滑及保护,促进创伤愈合。
透明质酸(HA):无硫酸化、无蛋白质共价结合,以游离或非共价复合体存在。细菌也可产生。二糖单位:GlcUAβ(1→3)GlcNAc。HA处于间隙时产生膨压,同时羧基吸引阳离子增加渗透压,使得结缔组织具抗压和弹性。HA吸水膨胀外基质利于细胞迁移。
硫酸软骨素(CS):二糖单位:GlcUAβ(1→3)GalNAc。硫化部位GalNAc的C4和C6位。体内以蛋白聚糖集体形式存于软骨、肌腱、韧带及主动脉等组织基质中。硫酸皮肤素(DS,软骨素B),优势单位IdoUAβ(1→3)GalNAc,少数为GlcUAβ(1→3)GalNAc。硫化部位IdoUA的C2位。DS是CS的一种修饰形式。作为结缔组织大分子存于皮肤、血管壁及心瓣膜。
硫酸角质素(KS):二糖单位:Galβ(1→4)GlcNAc。二糖单位β(1→3)连接。KS为糖胺聚糖中唯一不含糖醛酸单体的杂多糖。硫化部位Gal或/和GlcNAc的C6位。
硫酸乙酰肝素/硫酸类肝素(HS):具相同主链结构,分布于肺、动脉管等细胞表面,具抗凝活性,低于肝素。二糖单位:GlcUAβ(1→4)GlcNAc,GlcNAc上C2位具N-硫酸基修饰,硫化部位GlcNAc的C6位。肝素(Hp)存于肺、肝、皮肤及其它结缔组织的肥大细胞中,天然抗凝剂。二糖单位:IdoUAα(1→4)GlcNAc,GlcNAc上C2位具N-硫酸基修饰,硫化部位IdoUA的C2位或/和GlcNAc的C6位。
1.1.7.2. 蛋白聚糖¶
蛋白聚糖(PG),一类特殊糖蛋白,由一条或多条糖胺聚糖和一个核心蛋白共价连接。分布于细胞外基质、表面及胞内分泌颗粒。
核心蛋白,与糖胺聚糖共价结合的多肽链。多数核心蛋白具不同结构域,均含有相应糖胺聚糖结合结构域,某些蛋白聚糖通过核心蛋白特定结构域锚定于细胞表面或外基质大分子上,有些核心蛋白具特异相互作用结构域。
除透明质酸外,所有糖胺聚糖的延伸均在与核心蛋白共价连接的连接区(寡糖链)上进行。
大分子聚集型胞外基质蛋白聚糖,小分子富含亮氨酸胞外基质蛋白聚糖,跨膜胞内蛋白聚糖。
蛋白聚糖聚集体,以HA分子为主,经连接蛋白与核心蛋白相连,核心蛋白上分布由KS链及CS链。可聚蛋白聚糖对维持软骨形态及功能具重要意义。
1.1.8. 糖链结构分析¶
1.1.8.1. 一般步骤¶
糖蛋白的分离纯化:N-连接:PNG酶F、肼解。O-连接:O-糖苷酶、NaOH溶液β消去
释放完整聚糖
聚糖的分离纯化
聚糖纯度鉴定及相对分子质量测定
单糖组成测定
完整糖链序列测定。
1.1.8.2. 测定方法¶
化学方法:高碘酸氧化、甲基化分析、寡糖顺序降解。
酶学方法:
外切糖苷酶:非还原端,糖基、糖苷键专一性,提供单糖残基组成、顺序及糖苷键构型
内切糖苷酶:水解糖链内部糖苷键,释放糖链片段,从肽链上释放完整聚糖链
仪器测定:红外光谱(IR)、质谱(MS)、核磁共振(NMR)等。
1.1.9. 糖酵解¶
机体主要供能物质为ATP,ATP形成主要经Glc彻底氧化为CO2和水时释放大量自由能形成,或经Glc无氧条件下降解为丙酮酸仅产生2ATP。
糖酵解,无氧条件下葡萄糖分解为2分子丙酮酸并供能。是葡萄糖分解代谢共同途径。意义在于不需氧下产生ATP供能。
中间产物磷酸化:负电性磷酸基团使中间产物具极性而不易跨膜失散,对酶而言磷酸基团起信号作用利于与酶结合,磷酸基团经酵解后形成ATP末端磷酸基团利于保存能量。
糖酵解从Glc起至丙酮酸共10步反应:
前5步为准备阶段,葡萄糖经磷酸化、异构化裂解为三碳糖,裂解需要2ATP每己糖分子;
后5步为产生ATP的放能阶段,磷酸三碳糖转为丙酮酸,产生2ATP每三碳糖。
1.1.9.1. 准备阶段¶
葡萄糖磷酸化为G6P,由己糖激酶(Mg2+)催化将ATPγ-磷酸基团转移至Glc上;反应不可逆。利于葡萄糖的活化及防止葡萄糖跨膜。
反应时,ATP需与Mg2+形成复合物,未形成复合物的ATP对己糖激酶具强竞争性抑制作用。
己糖激酶受G6P、ADP变构抑制;葡萄糖激酶(肝)不受G6P抑制。 表 1.1.3
类型 |
部位 |
|
|---|---|---|
Ⅰ |
脑、肾 |
|
Ⅱ |
骨骼、心肌 |
|
Ⅲ |
肝、肺 |
|
Ⅳ |
肝 |
葡萄糖激酶受胰岛素诱导 |
G6P异构化为F6P,由磷酸葡萄糖异构酶催化,反应可逆。磷酸葡萄糖异构酶具绝对的底物专一性和立体专一性。6PG、E4P、S7P等皆为其竞争性抑制剂,也是戊糖磷酸途径的中间物。
F6P磷酸化为F-1,6-2P,由磷酸果糖激酶(Mg2+)催化,消耗1ATP,反应不可逆。
磷酸果糖激酶(表 1.1.4)为变构酶,催化效率低,糖酵解途径限速酶,哺乳类糖酵解调控关键酶;受高浓度ATP抑制,高H+浓度具抑制作用。抑制通过阻止酵解途径继续进行,防止乳酸积累、血液pH下降避免酸中毒。
类型 |
敏感抑制物 |
|---|---|
A |
磷酸肌酸、柠檬酸、无机磷酸 |
B |
BPG |
C |
AMP |
F-1,6-2P裂解为GAP及DHAP,由醛缩酶(表 1.1.5)催化,反应可逆。
类型 |
部位 |
|
|---|---|---|
ⅠA |
肌肉 |
|
ⅠB |
肝脏 |
|
ⅠC |
脑 |
|
Ⅱ |
细菌、酵母、真菌、藻类 |
含2价金属离子,Zn2+,Ca2+,Fe2+,K+ |
DHAP异构为GAP,由丙糖磷酸异构酶催化,以Glu游离羧基与底物结合,反应极迅速,可逆。
只有甘油醛-3-磷酸(GAP)继续进入糖酵解途径,二羟丙酮(DHAP)需转变为GAP才能继续进入途径。
准备阶段中,每分子葡萄糖进入,共消耗2ATP。
1.1.9.2. 放能阶段¶
GAP氧化为1,3-BPG,由GAPDH催化,NAD+及Pi参加,产生NADH及H+。砷酸盐结构及反应与Pi相似,在砷酸盐存在下,酵解可进行但无高能磷酸键形成,GAP氧化释放的能量未被磷酸化偶联;砷酸盐起解偶联作用,解除氧化与磷酸化的偶联。
1,3-BPG转移高能磷酸基团形成ATP,由PGK(Mg2+)催化,高效放能反应。生成等量的3-PG及ATP。
3-PG转变为2-PG,由磷酸甘油酸变位酶催化。变位酶指催化分子内化学基团移位的酶。中间物2,3-BPG,在红细胞中起调节剂作用,稳定脱氧血红蛋白降低氧亲和力;是二磷酸甘油酸变位酶强竞争性抑制剂。2,3-BPG的合成降解是糖酵解途径的一个短支路。
2-PG脱水为磷酸烯醇式丙酮酸,由烯醇化酶催化。氟化物是烯醇化酶的强抑制剂,氟与镁及Pi形成复合物替代Mg2+使酶失活。
磷酸烯醇式丙酮酸转变为丙酮酸,由丙酮酸激酶催化,产生1ATP。
丙酮酸激酶:是糖酵解途径中一个重要变构调节酶,活性需2价阳离子参与(Mg2+、Mn2+)
抑制作用:ATP、长链脂肪酸、乙酰CoA、Ala
激活作用:F-1,6-2P、磷酸烯醇式丙酮酸
放能阶段,每分子GAP进入能生成2ATP。1葡萄糖进入糖酵解至丙酮酸时,共消耗2ATP,产生4ATP,净获得2ATP。
1.1.9.3. 丙酮酸去路¶
无氧条件下,丙酮酸可生成乳酸或是生成乙醇。
生成乳酸,由乳酸脱氢酶(LDH)催化,需HADH;生成乳酸及NAD+。NAD+是GAP继续氧化的基础。
哺乳类LDH具M型(多则丙酮酸亲和力高)及H型(多则丙酮酸亲和力低)亚基,构成5种酶,心肌中的H4型对丙酮酸亲和力最低,利于丙酮酸脱氢酶催化朝有氧代谢进行。
机体内LDH同工酶比例可作为心肌、肝脏等疾病的重要指标。
生成乙醇,过程分两阶段,先由丙酮酸脱羧形成乙醛和CO2,再由乙醛与NADH及H+还原成乙醇同时产生NAD+。
第一步由丙酮酸脱羧酶催化,辅酶TPP,不存在于动物细胞中
第二步由乙醇脱氢酶(ADH)催化
1.1.9.4. 调节¶
糖酵解途径中,己糖激酶、磷酸果糖激酶及丙酮酸激酶催化的反应都不可逆。活性受变构效应物可逆结合及酶共价修饰调节。
磷酸果糖激酶是关键酶、限速酶,受高浓度ATP抑制,柠檬酸通过加强ATP抑制效应而抑制其活性。
F-2,6-2P是磷酸果糖激酶强激动剂,一种变构激活剂;提高果糖激酶与F6P的亲和力降低ATP抑制。
备注
葡萄糖缺乏时,胰高血糖素启动cAMP级联效应,双功酶(PFK2/FBPase2)磷酸化,酶共价修饰激活FBPase2、抑制PFK2,F-2,6-2P减少。葡萄糖过剩时,磷酸基团从酶上脱落,F-2,6-2P含量升高,糖酵解加速。
己糖激酶,受G6P抑制,非限速酶原因:G6P还可转变为糖原及参与戊糖磷酸途径。
丙酮酸激酶,受F-1,6-2P激活、ATP抑制、低Glc浓度可磷酸化而活性降低、Ala变构抑制。
1.1.9.5. 其它六碳糖进入途径¶
果糖:
肌肉中:由己糖激酶催化为F6P
肝中:经果糖激酶催化消耗1ATP成F1P,经果糖-1-磷酸醛缩酶(B型醛缩酶)催化为甘油醛及二羟丙酮磷酸,甘油醛经甘油醛激酶催化消耗1ATP成GAP,或在醇脱氢酶、甘油激酶及甘油磷酸脱氢酶催化下最终形成二羟丙酮磷酸消耗NADH、1ATP及NAD+
果糖不耐症,肝中缺乏B型醛缩酶造成F1P积累、Pi大量消耗至耗竭,ATP浓度降低,糖酵解大量产生乳酸。
半乳糖:
经半乳糖激酶催化消耗1ATP成Gal-1-P,经尿苷酰转移酶催化为UDP-Gal,经UDP-Gal4差向异构酶催化以NAD+为辅酶成UDP-Glc,经UDP-Glc焦磷酸化酶催化为G1P,经磷酸葡萄糖变位酶催化为G6P。
半乳糖血症,缺乏Gal1尿苷酰转移酶,半乳糖积累,使晶状体半乳糖含量提高并还原为半乳糖醇,最终形成白内障;严重引起生长停滞、智力迟钝、肝损伤甚至死亡。
甘露糖:经己糖激酶催化为Man6P,经磷酸甘露糖异构酶催化为F6P。
1.1.10. 柠檬酸循环¶
在细胞线粒体中进行,丙酮酸经TCA循环脱羧、脱氢,羧基成CO2、氢随NAD+、FAD进入电子传递链,经氧化磷酸化形成水并合成ATP。 TCA循环除丙酮酸氧化外也是脂肪酸、氨基酸氧化分解的共同途径;中间产物是众多合成前体;是两用途径。
1.1.10.1. 准备阶段¶
丙酮酸进入TCA循环需先转变为乙酰CoA。
丙酮酸脱羧,由E1催化为羟乙基-TPP;羟乙基氧化为乙酰基后转移至E2上。乙酰基转移至CoA上形成乙酰CoA,E2成还原型。在氧化型E3作用下再生氧化型E2;NAD+下再生氧化型E3。 表 1.1.6
酶系 |
缩写 |
辅基 |
反应 |
|---|---|---|---|
丙酮酸脱氢酶组分 |
E1 |
TPP |
丙酮酸氧化脱羧 |
二氢硫辛酰转乙酰基酶 |
E2 |
硫辛酰胺 |
乙酰基转移至CoA |
二氢硫辛酸脱氢酶 |
E3 |
FAD |
还原型硫辛酰胺转为氧化型 |
亚砷酸盐、有机砷化物与E2辅基硫辛酰胺巯基共价结合,成无催化能力砷化物(对α-酮戊二酸脱氢酶复合体具一致抑制作用)。
乙酰CoA既是柠檬酸循环入口物质,亦是脂类合成起始物。
调控关键:在于对丙酮酸脱氢酶复合体活性调控。
产物控制:乙酰CoA是E2竞争性抑制剂,NADH是E3竞争性抑制剂;
磷酸化状态的调控:E1磷酸化/去磷酸化是复合体失活/激活的重要方式。
1.1.10.2. 反应机制¶
从碳骨架层面看,呈现4666654444的变化。
草酰乙酸与乙酰CoA缩合成柠檬酸,由柠檬酸合酶催化。
柠檬酸合酶:TCA循环限速酶,属调控酶
活性受ATP、NADH、琥珀酰CoA、脂酰CoA等抑制。
抑制剂:氟乙酸(致死性合成反应)、丙酮基CoA。
柠檬酸异构为异柠檬酸,由乌头酸酶催化。
异柠檬酸氧化为α-酮戊二酸,由异柠檬酸脱氢酶催化脱羧,产生1CO2、1NADH。
异柠檬酸脱氢酶为变构调节酶,活性受ADP变构激活、受NADH、ATP变构抑制。
NAD+型:仅存于线粒体;NADP+型:线粒体及胞浆
α-酮戊二酸氧化为琥珀酰CoA,由α-酮戊二酸脱氢酶催化脱羧,产生1CO2、1NADH。
α-酮戊二酸氧化释放能量:驱使NAD+还原、促使反应向氧化方向进行、能量以琥珀酰CoA高能硫酯键保存。α-酮戊二酸脱氢酶受琥珀酰CoA、NADH及高能荷抑制;与丙酮酸脱氢酶类似,但不受磷酸化、去磷酸化共价修饰调节。
琥珀酰CoA转化为琥珀酸,由琥珀酰CoA合成酶催化,产生1高能键(哺乳类GTP,植物微生物ATP)。
琥珀酸脱氢为延胡索酸,由琥珀酸脱氢酶(以FAD作电子受体;具严格立体专一,嵌入线粒体内膜;含Fe-S聚簇,位于电子传递链上)催化,产生1FADH2。
延胡索酸水合为苹果酸,由延胡索酸酶(具严格立体专一,仅产生L-苹果酸)催化,消耗1H2O。
苹果酸脱氢为草酰乙酸,由苹果酸脱氢酶(辅酶NAD+)催化。ΔG>0,由草酰乙酸与乙酰CoA缩合形成的草酰乙酸消耗驱动反应。
TCA每周期共产生3NADH、1FADH2、1GTP、2CO2,消耗1乙酰CoA、2H2O。可共产生10ATP。
1.1.10.3. 调控¶
TCA循环系统内调节:
关键酶:柠檬酸合酶、异柠檬酸脱氢酶、α-酮戊二酸脱氢酶。
关键底物:乙酰CoA、草酰乙酸、NADH。
通常柠檬酸利用速率高于合成速率,柠檬酸是草酰乙酸的竞争性抑制剂。
ATP、ADP、Ca2+对TCA循环的调节:
ADP为异柠檬酸脱氢酶变构激活剂;
Ca2+具激活作用。
1.1.10.4. TCA双重作用¶
TCA具分解代谢及合成代谢的双重作用;中间物可参与糖异生、脂类合成、氨基酸合成、卟啉类合成等途径。
填补反应,对TCA中间物具补充作用的反应。如丙酮酸经丙酮酸羧化酶催化为草酰乙酸;乙酰CoA是丙酮酸羧化酶激动剂,最后可提高TCA催化速度。
降解途径产生TCA中间物的,奇数脂肪酸氧化;Ile、Met、Val分解产生琥珀酰CoA;脱氨基及转氨基作用产生α-酮戊二酸、草酰乙酸。反应皆可逆取决代谢需要。
TCA是代谢的中心,产生的NADH、FADH2经电子传递链、氧化磷酸化产生ATP。
1.1.11. 戊糖磷酸途径¶
戊糖磷酸途径为糖代谢第二条重要途径,在细胞溶胶种进行,广泛存于动植物细胞种。
核心反应:
总反应:
1.1.11.1. 氧化阶段¶
六碳糖脱羧形成五碳糖并使NADP+还原为NADPH。
G6P在G6P脱氢酶以NADP+为辅酶催化形成6磷酸葡萄糖酸δ内酯;实质不可逆,重要调控点。
6磷酸葡萄糖酸δ内酯在专一内酯酶催化水解成GlcA6P。
GlcA6P在GlcA6P脱氢酶催化成核酮糖5磷酸,以NADP+为电子受体。
1.1.11.2. 非氧化阶段¶
此阶段反应皆可逆。
核酮糖5磷酸由其异构酶经烯二醇中间物异构化Rib5P。与糖酵解G6P转化为F6P、DHAP转化为GAP同属酮醛异构反应,均经烯二醇中间物。
核酮糖5磷酸由其差向异构酶催化为木酮糖5磷酸。
备注
特别意义:转酮酶底物酮糖C3羟基需与木酮糖一致才具作用。
木酮糖5磷酸与Rib5P由转酮酶催化为GAP与景天庚酮糖7磷酸。
GAP与景天庚酮糖7磷酸由转醛酶催化为F6P与赤藓糖4磷酸。
木酮糖5磷酸与赤藓糖4磷酸由转酮酶催化为GAP与F6P。
F6P由磷酸葡萄糖异构酶催化为G6P。
1.1.11.3. 调控¶
G6P脱氢为限速反应,最重要调控因子为NADP+水平;
NADP+与NADPH竞争性抑制G6P脱氢酶及GlcA6P脱氢酶活性。
途径中G6P去路受NADPH、Rib5P、ATP的调节。可分三种情况:
Rib5P需求远大于NADPH需求:常于细胞分裂期。大量G6P经糖酵解转为F6P、GAP,再由转酮酶、转醛酶将2F6P、1GAP经方向途径成3Rib5P。
\(5G6P + ATP → 6Rib5P + H^+ + ADP\)。
Rib5P需求与NADPH需求平衡:此时氧化阶段处优势。反应为核心反应。
NADPH需求远大于Rib5P需求:G6P被彻底氧化;脂肪组织中戊糖磷酸途径比肌肉中更活跃。
促进以下反应:氧化阶段形成2NADPH、1Rib5P;Rib5P由转酮酶、转醛酶催化为F6P、GAP;F6P、GAP经糖异生为G6P。
\(G6P + 12NADP^+ + 7H_2O → 12NADPH + 12H^+ + 6CO_2 + Pi\)。
1.1.11.4. 意义¶
产生还原力(NADPH)的主要途径
细胞内不同结构糖分子的重要来源,为各单糖相互转化提供条件
缺乏G6P脱氢酶,使红细胞NADPH浓度低,易得贫血症
对具氧化性药物如奎宁、磺胺类、阿司匹林等过敏,因NADPH缺乏而使细胞易破裂,造成溶血性贫血
1.1.12. 糖异生¶
糖异生,葡萄糖异生作用,以非糖物质为前体合成葡萄糖,如乳酸、丙酮酸、丙酸、甘油及氨基酸等。
糖异生不是糖酵解的直接逆反应(表 1.1.7)
糖酵解中的三步反应为不可逆反应:
己糖激酶催化的Glc与ATP形成G6P与ADP
磷酸果糖激酶催化的F6P与ATP形成F-1,6-2P与ADP
丙酮酸激酶催化的磷酸烯醇式丙酮酸与ADP形成丙酮酸与ATP
糖酵解 |
糖异生 |
|---|---|
己糖激酶 |
葡萄糖6磷酸酶 |
磷酸果糖激酶 |
果糖-1,6-二磷酸酶 |
丙酮酸激酶 |
|
1.1.12.1. 途径¶
丙酮酸经草酰乙酸形成磷酸烯醇式丙酮酸:
丙酮酸由丙酮酸羧化酶催化消耗1ATP形成草酰乙酸,草酰乙酸由PEPCK催化为磷酸烯醇式丙酮酸。
\(丙酮酸 + ATP + GTP + H_2O → 磷酸烯醇式丙酮酸 + ADP + GDP + 2H^+ + Pi\)
F-1,6-2P由F-1,6-2P酶催化水解为F6P
\(F-1,6-2P + H_2O → F6P + Pi\)
G6P由G6P酶催化水解为Glc
G6P酶存于sER,活性需一Ca2+结合稳定蛋白协同作用;反应前G6P需转移至ER内,形成的Glc、Pi经不同转运途径至胞浆。
\(G6P + H_2O → Glc + Pi\)
肝、肠、肾细胞由G6P形成的Glc进入血液维持血糖。脑、肌肉无G6P酶;肝中糖异生主要为骨骼肌代谢产物乳酸、丙氨酸。
由丙酮酸形成葡萄糖:消耗4ATP、2GTP共6个高能磷酸键。
\(2丙酮酸 + 4ATP + 2GTP + 2NADH + 6H_2O → 葡萄糖 + 4ADP + 2GTP + 2NAD^+ + 2H^+ + Pi\)
1.1.12.2. 调节¶
磷酸果糖激酶、果糖-1,6-二磷酸酶的调节。
丙酮酸激酶、丙酮酸羧化酶、磷酸烯醇式丙酮酸羧激酶间的调节。
1.1.12.3. 乳酸再利用¶
肌肉细胞内乳酸进入血液流至肝脏,经糖异生为葡萄糖,随血液至肌肉、脑供给代;可立氏循环。
1.1.12.4. 葡萄糖转运体¶
葡萄糖出入细胞质膜非经简单扩散,依靠葡萄糖运载蛋白(表 1.1.8)。
类型 |
位置 |
功能 |
|---|---|---|
GLUT1、GLUT3 |
哺乳类细胞 |
基本Glc摄取 |
GLUT2 |
肝、胰β细胞 |
高Km |
GLUT4 |
肌肉、脂肪细胞 |
饱食时胰岛素促增量 |
GLUT5 |
小肠 |
Na+/Glc共转运蛋白 |
GLUT7 |
ER |
G6P进入ER |
1.1.13. 乙醛酸途径¶
此途径仅存于植物、微生物中
催化酶即存于线粒体,也存于植物膜特有的乙醛酸循环体(尤其是特有酶:异柠檬酸裂合酶、苹果合酶)。
\(2乙酰CoA + 2NAD^+ + FAD → 草酰乙酸 + 2CoASH + 2NADH + FADH_2 + 2H^+\)
1.1.14. 糖的其它代谢途径¶
糖苷键的形成需供能,主要以单糖活化的NDP-糖形式提供。
1.1.14.1. 乳糖合成及分解¶
乳糖:
由半乳糖和葡萄糖以β-糖苷键相连,合成前需活化的半乳糖前体:UDP-Gal;
由Gal1P尿苷酰转移酶催化:\(UDP-Glc + Gal1P → Glc1P + UDP-Gal\)
乳糖合酶由半乳糖基转移酶(催化UDP-Gal转移至GlcNAc)及α乳清蛋白(使Glc为Gal受体)构成。
乳糖于消化道由乳糖酶催化分解,微生物由β-半乳糖苷酶催化。
β-半乳糖苷酶为可诱导酶,相伴合成的有半乳糖通透酶、硫代半乳糖苷转乙酰基酶,E.coli中天然诱导物为别乳糖(IPTG、蓝白斑筛选)。
乳糖不耐症,乳糖于小肠产生强渗透效应使液体流向小肠、于大肠由细菌转为有毒物产生各症状。
1.1.14.2. 糖蛋白糖链合成及分解¶
糖蛋白:
N-糖链合成与肽链合成同步进行
O-糖链合成为肽链合成后对肽链逐个糖基修饰加工
合成由糖基转移酶催化,对供受体具严格专一性;首个糖基受体常为肽链特定位点氨基酸残基(Asn、Ser、Thr、Tyr、Hyp)
N-糖链水解由外侧肽链水解后再水解,常先水解为Fuc、后为β1,4-糖苷键连接的GlcNAc;肽链糖链分开后分别水解。
O-糖链水解依据糖链密集度,肽链、糖链水解次序不同或同时水解。
1.1.15. 糖原分解与合成¶
糖原,动物细胞最易动员的贮存葡萄糖,主要由肝、肌肉贮存;保证机体最需能量供应的脑、肌肉活动,保证不间断维持血糖平衡。
糖原分子90%降解为G1P,其余为葡萄糖。糖原以颗粒存于胞浆中,颗粒含合成降解酶及调节蛋白,利于贮存及动员。
肌肉动员糖原远快于脂肪,脂肪酸残基无氧条件下不能分解代谢,动物无法将脂肪酸转化为葡萄糖前体。
1.1.15.1. 降解¶
经磷酸解脱去Glc,由糖原磷酸化酶(辅酶:磷酸吡哆醛)催化,催化1→4糖苷键即直链Glc
\(糖原(n个Glc)+Pi→糖原(n-1个Glc)+G1P\)
糖原降解
采用磷酸解,使得降解的Glc带有磷酸基团,不需能量便可转变为G6P
水解则需1ATP转为G6P
同时生成的G1P以解离形式存在不易扩散至胞外
糖原经过糖原磷酸化酶催化至α(1→6)糖苷键前4个残基时暂停,后续降解需糖原脱支酶(含糖基转移酶)与糖原磷酸化酶协同作用。
生成的G1P经磷酸葡萄糖变位酶催化为G6P。
葡糖6磷酸酶,专门水解G6P,存于肝中,维持血糖平衡。
1.1.15.2. 合成¶
\(G1P + UTP → UDPG + PPi\),由UDP葡萄糖焦磷酸化酶催化。
糖原合酶催化将UDPG中Glc转至非还原端处,仅形成1→4糖苷键。
糖原分支酶,形成α(1→6)糖苷键,分支增加可溶性、非还原末端数,提高分解合成效率。
1.1.15.3. 调控¶
糖原合酶、糖原磷酸化酶受效应物别构调控,如ATP、G6P、AMP等。
糖原磷酸化酶:
AMP对骨骼肌磷酸化酶的别构调控,高AMP使酶b由T构象转为R,经激酶磷酸化后酶b转为酶a具活性。
其激酶的调控受cAMP激活的PKA活化和Ca2+激活。
神经冲动引起Ca2+释放,使糖原降解与收缩关联。
糖原合酶:磷酸化时无活性,非磷酸化时具活性。
肝中对葡萄糖敏感依赖:
Ser磷酸化和葡萄糖相关酶变构部位联系
PP1对磷酸化酶的钝化及激活糖原合酶
PP1与磷酸化酶a的结合阻止糖原合酶活性。
胰高血糖素刺激cAMP形成;胰岛素刺激糖原合成,去磷酸化解除糖原合酶抑制、磷酸化酶激酶及磷酸化酶a受抑。
Ⅰ型糖尿病为胰岛素依赖型,β细胞被自身免疫破坏;
Ⅱ型糖尿病为非胰岛素依赖,缺乏胰岛素受体。
糖原积累症,因缺失糖原代谢过程中某种酶而引起。
1.1.16. 光合作用¶
含光合色素的植物细胞及细菌在日光下利用无机物合成有机物,并释放氧或其它物质的过程。
光合作用发生于膜上,原核细胞中光合膜充满细胞,真核细胞中位于叶绿体。
叶绿体是内膜系统组织形式,类囊体膜组织成片层即类囊体小泡,垛叠成基粒,基粒间由基质片层相连;形成膜间隙、基质、类囊体腔。
1.1.16.1. 1.17.1 光反应¶
叶绿素吸收光能:光能由辅助色素收集,传递至主要色素叶绿素a(细菌叶绿素)。
叶绿素是深绿色光合色素的总称,为含镁四吡咯衍生物;
在高等植物、藻类中存有5种:a(存于所有放氧生物)、b、c、d、e。
辅助色素即藻胆素、类胡罗卜素、除叶绿素a、细菌叶绿素外的其它叶绿素;扩展光吸收的范围。
叶绿素在膜上组织成光合单位:
一个光合单位由叶绿素分子及其它色素分子组成天线加一个具光化学反应性的叶绿素a二聚体。
量子产额指每吸收一量子后释放的氧分子数。
光量子命运:热损失(内转换)、光损失(荧光)、共振能传递(激子传递,将光子传递至光化学反应中心)、电子传递(能量转换,光能转为化学能)。
光驱动的电子流:
光合细菌只有一个光化学作用中心,P870,将激子经细菌脱镁叶绿素或细菌叶褐素传递至醌或是将电子经醌传递至Fe-S中心;
高等植物、藻类具两光系统:PSⅠ(P700,分布于基粒片层、基粒顶底部)、PSⅡ(P680,分布于基粒内部紧密堆叠区)。
光合磷酸化:与氧化磷酸化类似,各复合体位于类囊体膜、可解偶联、可阻断、ATP合成类似。
每形成1O2:即4e-从H2O至NADP+,约12H+从基质转移至类囊体腔(4个H+由放氧复合体腔内产生;8个H+由Cytb6f转运,2H+/e-),合成约3ATP。
非循环式光合磷酸化
循环式光合磷酸化,无O2释放及NADP+还原。图 1.1.1
总反应: \(2 H_2O + 8 hv + 2 NADP^+ + 8/3 ADP + 8/3 Pi → O_2 + 2 NADPH + 8/3 ADP + 3 H2O\)
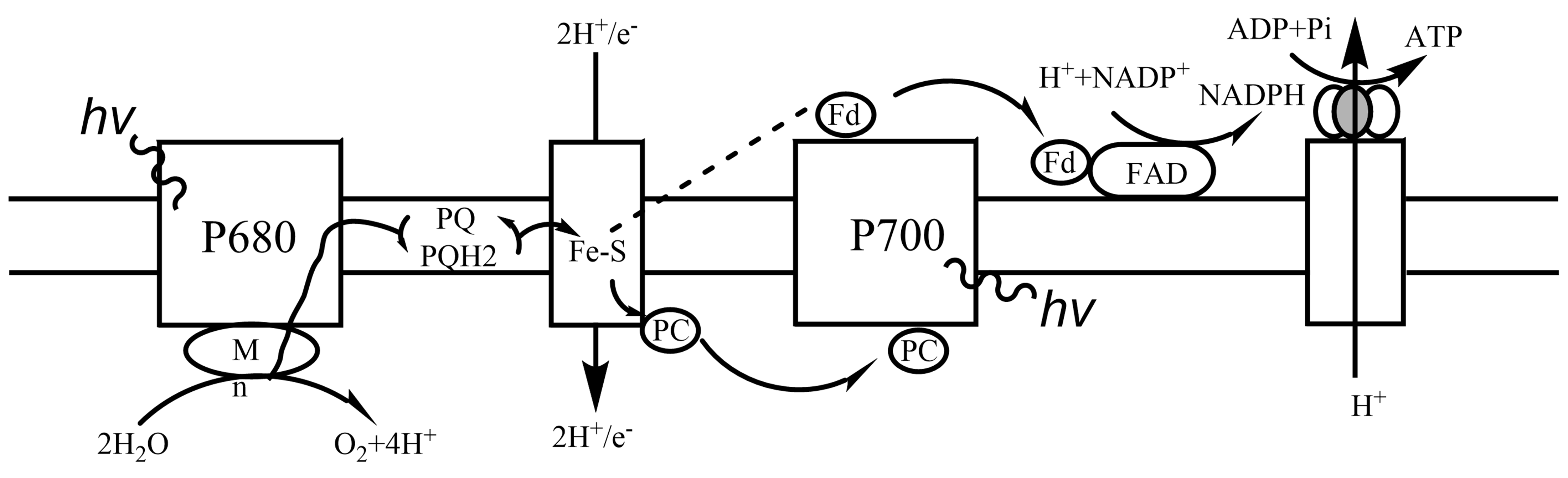
图 1.1.1 光合磷酸化¶
1.1.16.2. 暗反应¶
CO2固定:Rubisco羧化酶催化
\(6 CO_2 + 6 H_2O + 6 RuBP → 12 3-PG\)
还原:3-PG转化为G3P
\(12 3-PG + 12 ATP + 12 NADPH → 12 G3P + 12 ADP + 12 NADP^+ + 12 P\)
RuBP再生:从丙糖磷酸再生为RuBP。
CO2固定调节:基质pH变化、还原力的产生、Mg2+的外流。
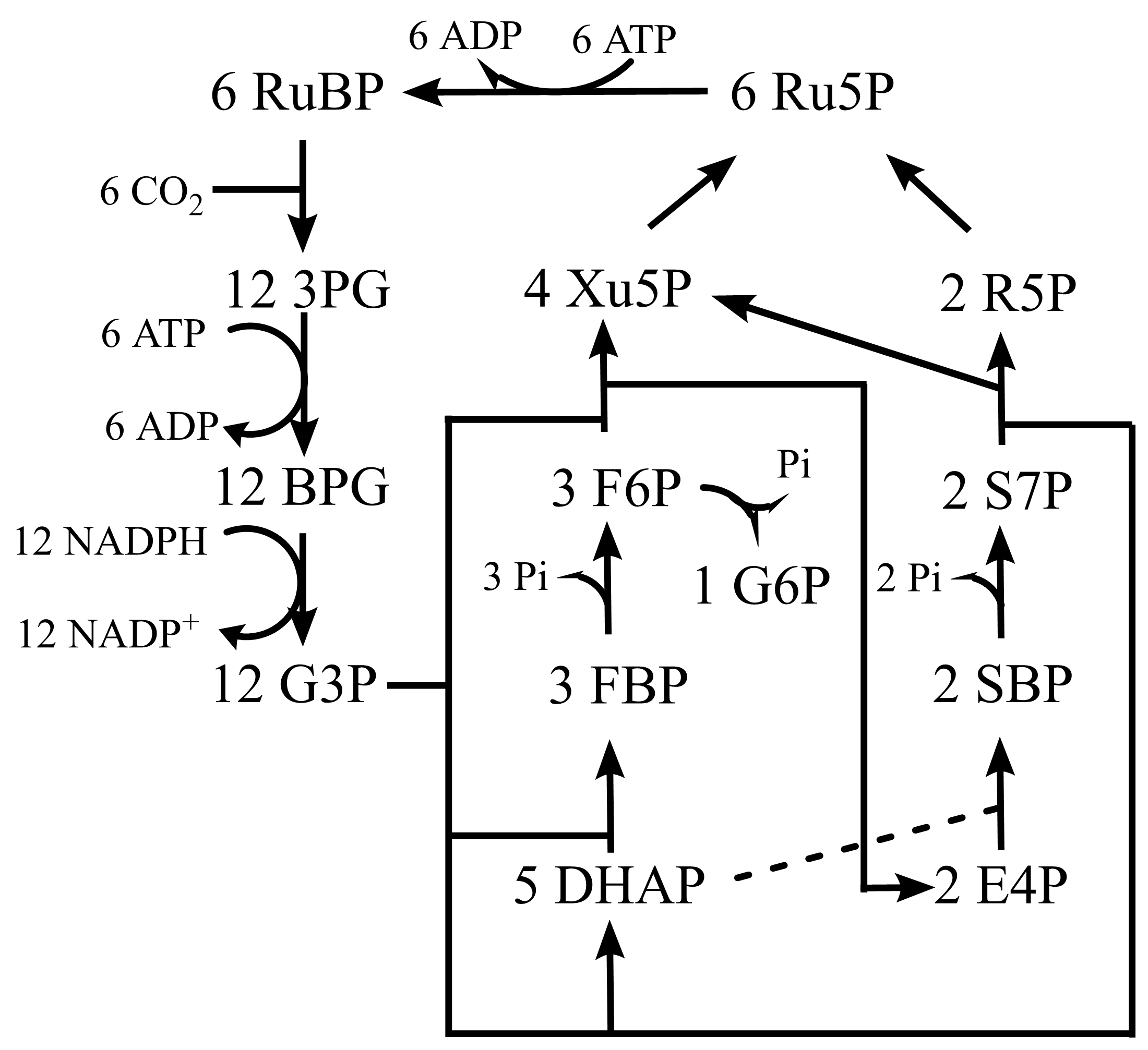
图 1.1.2 Calvin循环¶
1.1.16.3. 光呼吸及C4途径¶
光呼吸:Rubisco具羧化酶及加氧酶活性,温度升高对CO2亲和力下降
羧化酶:加氧酶活性比在室温大气平衡条件下为3:1;
加氧酶活性产生磷酸乙醇酸,经乙醇酸途径转为3-PG返回Calcin循环;
形成消耗O2产生CO2的过程。
C4途径:作为CO2收集、浓缩、转运系统;将从高氧叶片细胞转移至低氧内部细胞,减少光呼吸发生。
景天酸代谢(CAM):CO2吸收与固定在时间上分开。
CAM植物与C4植物中CO2均经C4途径进入Calvin循环,区别为:
CAM植物CO2最初羧化和糖合成发生于同一细胞的不同时间
C4植物则发生于同一时间的不同细胞。
1.1.16.4. 能量利用效率¶
CO2固定成己糖:
\(12 NADPH + 12 H^+ + 18 ATP + 6 CO_2 + 12 H_2O → C_6H_{12}O_6 + 12 NADP^+ + 18 ADP + 1 8Pi\)
需要合成1M己糖需12M NADPH、18M ATP;
按8Mhv产生8/3M ATP算,需要(48+6)Mhv,共9180KJ;
合成糖为2870KJ,效率31%。